Corsa ed Evoluzione della Specia Umana

L'anatomia umana si sarebbe sviluppata più di 2 Milioni di Anni fa sotto la
pressione e lo stimolo indotto da un 'attività di corsa, poiché
attività necessaria ai nostri antenati nel competere con gli altri
predatori per l'approvvigionamento di cibo nell'ambiente della
savana. Proprio la corsa, in particolare la corsa su lunghe
distanze, avrebbe stimolato la differenziazione dello scheletro e
dei complessi muscolari umani a partire dalle Australopitecine.
Questo il risultato, pubblicato il 18 Novembre 2004 sulla rivista
Nature, dei decennali studi del Biologo Dennis Bramble
dell'Università dello Utah e l'antropologo dell'Università di Harvard
Daniel Lieberman.
Premessa
I fossili relativi ai più antichi ominidi sono databili tra 6-3
Miliano di Anni (abbr. MA) , un periodo che va dal Tardo Miocene al Pliocene Medio.
Comunemente denominate Australopitecine arcaiche, questi fossili
rappresentano gli antenati più vicini alle Antropomorfe che
possediamo, forme ancora decisamente arcaiche, ma che già possiedono
caratteristiche derivate, prima fra tutte la locomozione bipede
abituale. I dati non sono certi a riguardo, essendo i complessi
fossili ancora scarsi e frammentari, ma informazioni a livello
dell'omero, della tibia, del femore e a livello craniale
indicherebbero che già questi arcaici ominidi si muovessero con
deambulazione bipodalica. un gruppo di resti trovato sulle colline
Tugen, in Kenya, relativo a porzioni di mandibole con denti sparsi e
frammenti degli arti inferiori, rivelerebbe che già l'Orrorin
tugenensis (tale è il nome di questo primo potenziale ominide) fosse
bipede, pur mantenendo un notevole adattamento all'arrampicata
arborea. Questi caratteri fondamentali sarebbero confermati anche
negli ominidi successivi, dai resti craniali relativi
all'Ardipithecus ramidus, che mettono in luce la posizione del
foramen magnum più avanzata rispetto alle Antropomorfe, così come ne
l'Australopithecus anamensis, il più antico rappresentante della
specie Australopithecus, dove caratteristiche a livello della tibia
attesterebbero il bipedismo. Questo carattere non si sarebbe perso nei
generi e nelle specie successive ma si sarebbe bensì affermato come
un carattere vantaggioso e quindi favorito dalla selezione
naturale, sino a rappresentare una caratteristica distintiva del
genere Homo. Gli Studi di Dennis Bramble e Daniel Lieberman
suggeriscono tuttavia che l'acquisizione della locomozione bipede di
per sé non rappresenti per l'uomo un deciso stacco evolutivo, un
carattere discriminante e stimolante cui ricondurre completamente
l'evoluzione anatomica di Homo habilis, e ancor più di Homo
ergaster. Le Australopitecine hanno camminato in posizione eretta
per almeno 2,5 milioni di anni, pur presentando ancora caratteristiche
fisiche ben lontane dall'uomo moderno: come può quindi il bipedismo
aver improvvisamente stimolato l'evoluzione del corpo verso
l'anatomia moderna? Questa la provocatoria e stimolante domanda
posta dai due studiosi americani.
Il significato evolutivo degli adattamenti anatomici in Homo spiegati attraverso la corsa
Generalmente Homo habilis viene considerato degno di
appartenenza al nostro stesso genere per caratteristiche a livello
craniale, quale la conformazione più umana della scatola cranica, con
gli occipitali arrotondati, e l'espansione delle aree frontali e
parietali, indici di una capacità cranica maggiore che si aggirava
attorno ai 600 cc. Ma non solo. Homo habilis è soprattutto il primo
ominide a sviluppare tratti comportamentali tipicamente umani,
l'occupazione ripetuta di una singola località, e la concreta
produzione di strumenti in pietra. Si tratta della più antica
testimonianza di una manipolazione volontaria e ripetuta dell'ambiente
esterno da parte di un animale. L'interpretazione dei choppers e
chopping tools, se lecito chiamarli così, è in realtà oggi argomento
di accesi dibattiti incentrati attorno alla loro presunta
funzionalità: o come nucleo, quindi esclusivamente come base per il
distaccamento di schegge, o come vero e proprio strumento di taglio,
incisione e percussione. Ciò che è importante mettere in evidenza, al
di là delle forti problematiche annesse allo studio delle industrie
relative al Paleolitico inferiore, è l'atteggiamento: l'aver
iniziato in modo sistematico a lasciar traccia, in ogni piano di
frequentazione, di ciottoli scheggiati e schegge, lavorate e non,
comunque utilizzate, da quanto l'analisi delle tracce d'uso rivela,
per tranciare, tagliare, raschiare, carne, legno, e cuoio.
(L'analisi delle tracce d'uso si basa sulla possibilità di
riconoscere lungo il margine funzionalmente attivo degli strumenti
tracce di lavorazione e di usura attraverso microscopio ottico a
luce riflessa, microscopio binoculare e microscopio a scansione,
potendo così, attraverso un confronto con le tracce e le usure
lasciate dalle attività sperimentali su manufatti litici moderni
fabbricati con il medesimo tipo di selce, determinare l'effettivo
utilizzo dello strumento.) Quale può essere stato il motore alla base
di un tale comportamento? Probabilmente una scoperta casuale, ma ciò
non toglie che successivamente a una semplice acquisizione
individuale si passi a una conoscenza comune, estesa non solo a
tutti gli elementi del singolo gruppo, ma anche agli altri gruppi di
ominidi e quindi a tutti gli appartenenti alla specie, in modo
relativamente uniforme, e senza che sia stato possibile identificare
sino ad adesso linee divergenti dalla tradizione Olduwaiana.
Quest'uniformità innanzitutto testimonia l'importanza
dell'apprendimento sociale come mezzo per trasmettere, anche
indirettamente, nuovi comportamenti utili alla sopravvivenza. Ogni
acquisizione umana, in questo contesto, deve infatti necessariamente
avere una ragione evolutiva, nascere con il fine di migliorare la
propria condizione e di presentarsi più competitivi nella lotta per la
sopravvivenza. Proprio lo stimolo base della conservazione di sé e
della propria specie rappresenterebbe il motore per l'acquisizione di
tali conoscenze e per la loro diffusione, in quanto necessarie, o
perlomeno utili, alla specie. Ed è in tale ottica che, secondo gli
studiosi americani, è effettivamente difficile spiegare lo sviluppo di
alcuni caratteri anatomici moderni attraverso la sola acquisizione
della locomozione bipodalica, anche perché in taluni casi non
interessati nella deambulazione e non funzionali a tale scopo. La
risposta viene quindi ricercata in un'attività naturale per l'uomo,
che è però stata spesso vista semplicemente come una conseguenza
logica della stazione eretta, la "corsa".
Bramble e Lieberman hanno ipotizzato che proprio la necessità di
correre abbia portato alla differenziazione della linea umana,
attribuendole una fondamentale importanza nella nostra storia
evolutiva. La selezione naturale avrebbe infatti favorito l'affermarsi
di quei caratteri anatomici che permettono di percorrere lunghe
distanze a corsa, necessitando quindi di una struttura ossea e
complessi muscolari che associno resistenza e potenza a stabilità ed
equilibrio. La deambulazione bipede, secondo tali recenti studi, non
figurerebbe quindi come un carattere così determinante nello
sviluppo dell'anatomia umana, bensì il bipedismo, sviluppatosi già
attorno a 4,5 MA in un habitat ancora parzialmente forestale, ben
prima dell'avvento dei primi uomini, non rappresenterebbe da solo
una caratteristica esclusiva e distintiva dell'uomo. Le
Australopitecine già camminavano bipedi pur presentando
caratteristiche fisiche decisamente distanti dalle proporzioni
odierne. Come spiegare quindi unicamente attraverso l'acquisizione
della postura eretta e della locomozione bipede tutti i cambiamenti
che distinguono Homo da Australopithecus che, rispetto a noi,
presenta gambe corte, avambracci più lunghi, spalle strette con la
cavità glenoidea leggermente inclinata verso l'alto e connessioni
muscolari tra spalle e il complesso testa-collo più estese e
robuste? Per arrivare alle loro conclusioni Bramble e Lieberman hanno
esaminato 26 tratti del corpo umano, di cui molti riscontrabili
anche nei reperti fossili relativi a Homo erectus e in parte a Homo
habilis, e solo alcuni di questi sono implicati nella deambulazione.
Caratteri anatomici necessari alla corsa
Fondamentalmente per praticare un'attività di corsa costante, il
corpo deve essere capace di assorbire l'impatto con il terreno,
assorbire quindi i colpi attraverso sistemi di dissipazione e
distribuzione dell'energia, deve essere stabile e in grado di
mantenersi in equilibrio, deve presentare una muscolatura
sufficientemente potente da poter prolungare il movimento nel tempo e
deve contemporaneamente essere capace di mantenere la temperatura
corporea costante per evitare il surriscaldamento.
Equilibrio e stabilità
Ai fini dell'equilibrio il nostro corpo innanzitutto presenta
una struttura craniale articolata sopra la colonna vertebrale, col
baricentro situato in posizione centrale. Questo permette di mantenere
la testa in posizione orizzontale con un minor dispendio di energie.
Contemporaneamente la porzione facciale si presenta più piatta
rispetto alle Australopitecine, con minor prognatismo, denti più
piccoli e un apparato masticatorio di minor peso e potenza. La parte
anteriore del cranio è controbilanciata dalla porzione occipitale,
distribuendo così equamente il carico. La corretta distribuzione del
peso è un carattere fondamentale soprattutto in attività di intenso
sforzo fisico quali la corsa. In aggiunta, il canale semicircolare
posteriore è, in Homo, decisamente più largo che in Pan o
Australopithecus, testimoniando probabilmente una maggiore
sensibilità alle scosse e ai sobbalzi che subisce la testa, che sono
potenzialmente maggiori e più intensi nella corsa che nella
camminata. Il torace umano presenta una struttura indipendente dalla
testa e le spalle hanno una posizione abbassata rispetto ad
Australopithecus e Pan: sono più larghe e l'unico muscolo che le
collega alla testa è il trapezio. Se infatti una struttura più
compatta, con spalle in posizione elevata e robuste connessioni
muscolari tra torace, collo e testa, è funzionale all'arrampicata
arborea, tale struttura non offre vantaggi nella locomozione bipede, ma
impedisce la rotazione del torace e degli arti superiori
indipendentemente dalla testa, movimento invece necessario nel
controbilanciare l'azione degli arti inferiori. La riduzione delle
dimensioni degli avambracci è fondamentale per ottimizzare la corsa
(anche oggi, individui con arti inferiori notevolmente più lunghi degli
arti superiori, risultano avvantaggiati in tale attività), riducendo
la potenza muscolare necessaria a flettere gli arti nel
bilanciamento. Il loro movimento deve richiedere uno sforzo
muscolare minimo, e allo stesso tempo compensare il movimento delle
gambe con un corretto andamento dei gomiti, che mai devono restare
fermi, per esempio in posizione arretrata, nel qual caso ci sarebbe
necessariamente la torsione del busto. Il tronco nella corsa deve
essere infatti portato avanti praticamente senza movimenti, con
l'asse sagittale in posizione verticale, senza che si abbia
"tilting", ossia, non deve aumentare l'inclinazione in avanti del
busto al momento dell'appoggio al terreno, per poi diminuire in fase
di spinta.
Ai fini della stabilità il corpo deve essere capace di assorbire
i contraccolpi, che nella corsa si fanno decisamente più forti. A
livello rachideo dorsale sono il legamento longitudinale anteriore e
il legamento longitudinale posteriore che ricoprono tale funzione.
Si tratta di fasce fibrose che si addossano rispettivamente alla
faccia anteriore e posteriore dei corpi vertebrali, dall'epistrofeo
sino alla parte superiore dell'osso sacro, aderendo fortemente ai
corpi delle vertebre, e più lassamente ai dischi vertebrali. Tali
legamenti sono riscontrabili in mammiferi specializzati nella corsa,
quali cani e cavalli, o in animali con teste di grosse dimensioni,
come elefanti, mentre non sono riscontrabili in Pan, e probabilmente
neppure in Australopithecus, non essendone rilevabile alcuna
traccia nei resti fossili. Le vertebre stesse già da Homo habilis
segnano una diminuzione in lunghezza, e un aumento del diametro
relativo all'aumento delle dimensioni dell'intero corpo, maggiore di
quanto non sia nei suoi antenati, permettendo alla schiena di
sopportare un maggior carico e di assorbire meglio gli sforzi.
Le connessioni tra la pelvi e la spina dorsale sono infine più
robuste e larghe, provvedendo a offrire maggiore stabilità, e la
pelvi nell'uomo, a differenza delle Australopitecine, presenta
inoltre rilievi ossei per gli attacchi dei muscoli glutei sulla
regione esterna. I glutei nell'uomo sono più ampi, più robusti, sono
muscoli critici durante la corsa connettendo il femore al tronco, e
in particolar modo è di rilievo il grande gluteo, che nell'uomo è
il più possente muscolo estensore dell'anca.
Robinson nel 1972 aveva già dimostrato che una persona colpita
da paralisi al grande gluteo poteva camminare senza gravi difficoltà,
ma a confermare ulteriormente le osservazioni di Bramble e
Lieberman contribuiscono le conclusioni a cui giunse nel 1972 V.
Basmajian. La sua scoperta rivelò che il grande gluteo nella
deambulazione su terreno pianeggiante ha una funzione esclusivamente
di controllo degli arti inferiori nel momento di massima estensione,
frenando lo slancio in avanti e conferendo all'arto stabilità nel
momento in cui tocca terra, e non ricopre nel cammino una funzione
propulsiva. Una normale camminata richiede quindi ben poco della
potenziale energia del grande gluteo, e questo vale anche per la sola
stazione eretta, mentre l'immenso potenziale del muscolo entra in
gioco quando l'individuo sale un pendio scosceso, si alza da
posizione seduta o corre, contribuendo alla stabilità del tronco e
offrendo la potenza necessaria a compiere il movimento. Considerando
che un'attività fisica per essere capace di stimolare alterazioni
anatomiche deve essere necessariamente un'attività costante,
continua e ripetuta più e più volte nel tempo, e prendendo in
considerazione l'ambiente stesso della savana, dove i primi uomini
si svilupparono, un ambiente prevalentemente pianeggiante, è
possibile ipotizzare che una grossa componente di responsabilità
nello sviluppo del grande gluteo sia da attribuire proprio alla
corsa.
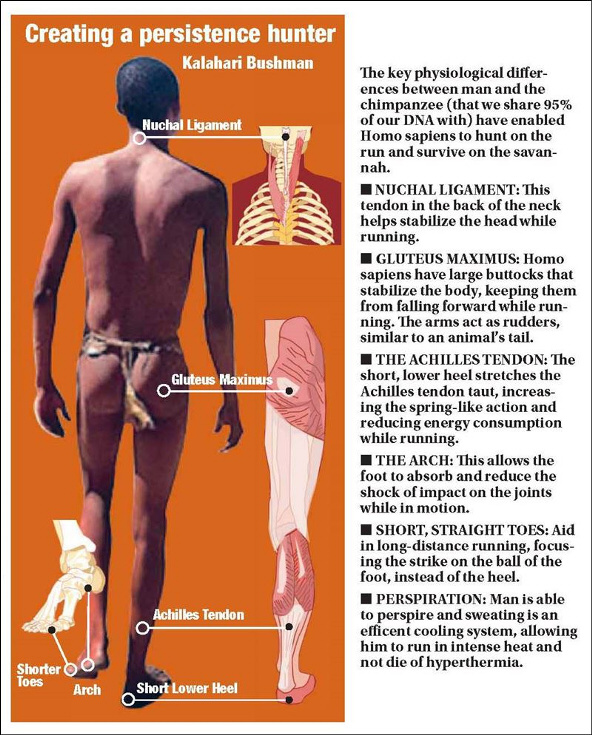
Termoregolazione
L'altezza raggiunta da H. ergaster, la cui media si aggirava
attorno ad 1,75 cm, comporta un aumento della superficie corporea e
permette quindi attraverso la sudorazione di dissipare meglio il
calore in attività che richiedono un estremo sforzo energetico. Allo
stesso tempo arti inferiori più lunghi, di cui non disponevano le
Australopitecine, permettevano una maggiore agilità e velocità.
Proprio nella corsa la lunghezza degli arti inferiori è importante
poiché l'aumento della velocità non si registra tanto con l'aumento
della frequenza dei passi, che comporta infatti anche un maggiore
dispendio di energie, ma con l'aumento della lunghezza di ogni singola
falcata. Inoltre fondamentale è lo sviluppo in H. ergaster di un
setto nasale vero e proprio, con narici rivolte verso il basso, che
può essere spiegato come risposta alla necessità di trattenere
l'umidità corporea durante momenti d'intensa attività fisica.
Meccanica della corsa e ruolo del piede nell'assorbimento e nella trasformazione dell'energia
Importantissimi sono poi gli adattamenti a livello del piede che
per sopportare l'impatto e i traumi sollecitati dalla deambulazione e
dalla corsa deve presentare una struttura robusta e allo stesso
tempo elastica. Ogni qualvolta che infatti il piede tocca il terreno
vi esercita delle forze e contemporaneamente ne subisce altre
uguali e contrarie, in relazione all'accumulo di energia elastica
all'impatto e al suo riutilizzo nella fase di spinta, per il
principio di conservazione dell'energia. Nella fase d'impatto la
forza reattiva agisce in direzione opposta al piede con effetto
frenante. I tendini del piede quindi si rilassano completamente
sotto l'azione gravitazionale, non opponendosi alla gravità ma
sfruttandola nell'adattare la pianta alla superficie d'appoggio.
Nella fase di stacco invece la forza reattiva determina lo
spostamento del corpo in avanti e il piede si oppone alla forza della
gravità, irrigidendosi (fondamentale a tale scopo è la migrazione
sottoastragalica del calcagno). Se però nella deambulazione le
problematiche energetiche sono risolte trasformando continuamente
l'energia potenziale in cinetica (con l'aumentare di una
proporzionalmente diminuisce l'altra), nella corsa invece i processi
di accumulo e trasformazione dell'energia sono più complessi,
sfruttando infatti in tal caso l'energia elastica del piede.
Analogamente a una palla che rimbalza, in cui lo spostamento in
avanti è dovuto all'energia elastica accumulata in seguito alla sua
deformazione nell'attimo in cui urta contro il suolo, nell'uomo
l'energia immagazzinata, proporzionale all'energia cinetica
sviluppatasi nella caduta, viene restituita nella componente
elastica (i muscoli), e negli elementi elastici in serie (i
tendini), durante la successiva spinta in avanti o in alto. In ogni
passo di corsa si individuano tre fasi: una fase di spinta, una di volo,
in cui il corpo è sospeso in aria, e una di arrivo al suolo. Il
lavoro che determina gli spostamenti del centro di gravità del corpo
viene compiuto essenzialmente nella fase di spinta. Da un lato
infatti si accelera il corpo, si aumenta dunque la sua energia
cinetica, dall'altro lo si innalza, aumentando la sua energia
potenziale. Nella fase di volo il baricentro del corpo raggiunge il
livello massimo e poi tende a scendere mentre la velocità rallenta,
se pur di poco, a causa della resistenza opposta dall'aria. Nella
fase d'impatto il centro di gravità si abbassa ulteriormente,
l'energia potenziale tocca il punto più basso, mentre il contatto
del piede con il terreno determina un rallentamento del corpo così
che anche l'energia cinetica arriva al valore minimo. Per mantenere
la velocità costante l'accelerazione che si verifica nella
successiva spinta dovrà necessariamente compensare la "frenata"
d'arrivo a terra e quella, di entità molto minore, dovuta alla
resistenza dell'aria. Per sviluppare quindi la forza necessaria alla
propulsione del corpo in avanti si sfrutta l'energia elastica
accumulata dai complessi mio-tendinei del piede e della gamba.
Fondamentali sono il tendine di Achille e la fascia plantare,
paragonabili a una molla che prima si contrae sotto l'azione di
forze e poi rilascia completamente l'energia immagazzinata. Il
tendine di Achille è la molla principale che accumula energia
elastica e il suo corretto funzionamento è tanto fondamentale per la
corsa che Bramble e Lieberman ritengono che da un punto di vista
evolutivo il suo sviluppo sia stato stimolato proprio da una
prolungata attività di corsa, mentre il suo coinvolgimento nella
deambulazione resterebbe relativamente scarso. La fascia plantare
invece ha un funzione fondamentale nel contribuire alla stabilità
dell'appoggio e nella trasmissione della forza dai muscoli del polpaccio
verso l'avampiede e la sua elasticità permette di risparmiare, per
mezzo della sua distensione, una notevole quantità di energie nella
corsa o nel salto. La loro importanza e il loro coinvolgimento
nell'attività fisica è testimoniato anche dalla frequenza con cui si
sviluppano negli atleti patologie relative a tali sistemi
muscolo-tendinei-ligamentosi, solitamente causate da carichi di lavoro
esagerati o incrementi troppo repentini, o da un errato appoggio del
piede durante l'attività sportiva Gli adattamenti che è possibile
rilevare dalle evidenze fossili di H. ergaster e H. sapiens includono
anche lunghi e "molleggiati" tendini, tra cui appunto il tendine di
Achille, mentre i reperti relativi ad Australopithecus suggeriscono
che le Australopitecine mancassero di un tendine di Achille ben
sviluppato, come così anche le attuali scimmie. L'arco plantare pare
essersi sviluppato solo parzialmente nelle Australopitecine, essendo
stato rilevato, dallo studio dei fossili di Sterkfontein e Hadar,
uno scafoide tarsale ben sviluppato e atto a sostenere più peso di
quanto non si registri nell'uomo, ma assai vicino alle proporzioni
di uno scimpanzé.
La struttura del piede nell'anatomia moderna si è notevolmente
differenziata rispetto ai nostri parenti più vicini, le Antropomorfe,
dovendo subire notevoli adattamenti legati soprattutto alla
stazione eretta costante, alla deambulazione e allo sviluppo di
movimenti sempre più complessi. La differenza più appariscente è
innanzitutto la divergenza dell'alluce, che passa da una funzione
prensile a una funzione di propulsione. Nell'uomo è strutturato
parallelamente alle altre dita, perdendo gran parte della mobilità
che possedeva. Ha prevalentemente il compito di liberare l'energia
accumulata, sfruttando la reazione della superficie d'appoggio nel
far progredire il corpo, e d'altro canto contribuisce a irrigidire
l'intera struttura del piede. La flessione dorsale delle dita
infatti, oltre ad aumentare l'ancoraggio al suolo nella spinta in
avanti, attiva anche un meccanismo ad argano, che rende il piede una
vera e propria barra di leva. Nella corsa la funzione delle dita e
dell'alluce sono ancor più evidenti, mancando spesso l'appoggio del
tallone, e venendo così scaricato improvvisamente tutto il peso
corporeo sull'avampiede. Essendo maggiore l'energia cinetica nella
caduta del corpo, deve essere maggiore anche la reazione, e quindi
l'energia elastica nella propulsione. L'alluce, che maggiormente si
fa carico del compito di spingere avanti il corpo, presenta quindi
una struttura ossea, relativamente alle Antropomorfe ed agli altri
Primati, molto più robusta rispetto alle altre dita. Riguardo alle
Australopitecine i dati sul piede sono ancora contrastanti e non
abbondanti. Nel 1979, Mary Leakey trovò dozzine di impronte (fig. 3)
in una località dell'Africa orientale chiamata Laetoli, in Tanzania.
orme che non si differenziano molto da quelle lasciate dai piedi di
essere umani odierni, ritrovate in strati costituiti da ceneri
vulcaniche solidificate 3,56 MA. Le impressioni appartengono a più
individui diversi, suggerendo a un'approfondita analisi un tallone
pronunciato, un arco longitudinale mediale sviluppato, con il peso
che poggia lateralmente, e un alluce lievemente divergente, di
dimensioni notevolmente maggiori rispetto alle altre dita. Queste
interpretazioni non sono in realtà uniformemente accettate, e ancor
oggi la discussione sulle impronte di Laetoli e sulla loro
appartenenza non è conclusa. Basti pensare che c'è chi ritiene che
siano impronte lasciate da esseri umani di anatomia moderna e chi
sostiene invece che siano state lasciate da forme ancestrali
contemporanee a Lucy. Ron Clarke nel 1998 scoprì uno scheletro
praticamente completo di Australopiteco in località Sterkfontein, in
Sud Africa. L'esemplare è stato datato a circa 3,7 milioni di anni
fa, come le impronte di Laetoli, ma Ron clarke ricostruì i piedi del
suo Australopiteco in termini decisamente più scimmieschi,
presentando tratti ancora arcaici. L'alluce è senza dubbio divergente,
allungato e proteso lateralmente e così pure le altre dita sono
decisamente allungate, più di quanto riscontreremmo nell'anatomia
moderna, ma comunque meno divergente di quanto non lo sia nelle
Scimmie Antropomorfe. L'interpretazione che fu data da Ron clarke e
dal paleoantropologo Phillip V. Tobias, leggeva la divergenza e
l'opponibilità dell'alluce come testimonianze di uno stile di vita
ancora parzialmente arboreo, associato però alla completa capacità di
progredire bipede ma, nonostante ciò, numerose restarono le voci di
dissenso, che vedevano l'Australopithecus africanus completamente
adattato alla locomozione su due piedi e alla vita terricola,
interpretando e facendo fede sulle evidenze a livello della spina
dorsale, dell'anca e del ginocchio. Ron Clarke continuò però nel
sostenere che se pur la divergenza dell'alluce in Australopithecus
afarensis era un carattere arcaico da legare a uno stile di vita ancora
parzialmente forestale, questo non escludeva l'adozione di una
locomozione bipede più o meno costante. Fondamentale a riguardo risultò
un esperimento portato avanti nel Boswell Wilkie Circus con due
scimpanzé, un maschio e una femmina, lasciati liberi di camminare su
sabbia bagnata. I risultati si rivelarono in accordo con le sue teorie.
La femmina camminava nervosamente, con insicurezza, irrigidendo la
pianta del piede, contraendo quindi le dita ed estendendo l'alluce;
invece il maschio era più sicuro e tendeva a camminare con l'alluce
in una posizione generalmente vicina alle altre dita, probabilmente
per una locomozione più comoda. La divergenza dell'alluce quindi di
per sé non rappresenta un ostacolo insormontabile nell'assumere la
stazione eretta e una deambulazione bipede, anzi l'esperimento
dimostra come in caso di necessità lo scimpanzé maschio riesca
addirittura ad adattarsi al terreno convergendo l'alluce verso le
altre dita per spostarsi più facilmente. Le impronte lasciate dallo
scimpanzé assomigliavano notevolmente alle impronte di Laetoli, e
dimostrano che una posizione dell'alluce perlopiù parallela alle
altre dita durante la locomozione bipede non implica necessariamente
lo sviluppo di una morfologia del piede vicina a quella umana, né
la perdita della capacità, se pur parziale, di opporlo. in
quest'ottica le impronte di Laetoli sarebbero potute anche essere
lasciate da un piede simile a quello dell'Australopithecus afarensis
StW573 ritrovato da Ron Clarke.
Inoltre Deloison suggerì che un'altra caratteristica
fondamentale delle impronte di Laetoli è l'assenza dell'impressione di
ogni singolo dito, a parte ovviamente l'alluce, come se fossero
contratte, proprio come le contrae Pongo quando procede bipede.
Nonostante però tutte le spiegazioni formulate e le interpretazioni
proposte, il mistero di Laetoli sembra essere destinato a fomentare
ancora a lungo disaccordo all'interno della comunità scientifica e non
solo. L'ipotesi avanzata da Bramble e Lieberman offre comunque
un'ulteriore chiave di lettura per le conoscenze finora accumulate
sulle Australopitecine e su Homo. i caratteri scheletrici relativi ad
Australopithecus rivelano infatti nell'insieme una morfologia
derivata rispetto alle Antropomorfe, in via di "umanizzazione", e
proprio la corsa potrebbe forse rappresentare lo stimolo che avrebbe
sempre più differenziato lo scheletro verso l'anatomia moderna. In
quest'ottica anche l'esperimento di Ron Clarke e le sue intuizioni
sulla possibile locomozione bipede di StW573, potrebbero suggerire
come per il raggiungimento della stazione eretta e della
deambulazione non sia stato necessario sviluppare tutti quei
caratteri "moderni" che oggi ci distinguono, ma si sia trattato di un
adattamento, se pur fondamentale, che ha apportato solo parziali
modifiche alla scheletro dello Australopitecine. Quindi proprio la
necessità di un'ulteriore specializzazione, la necessità di rendersi
competitivi nella corsa per la sopravvivenza e
nell'approvvigionamento del cibo avrebbe spinto l'uomo a
distinguersi e differenziarsi dai suoi antenati.
Funzione della corsa: dieta e caccia
Perché la corsa si rese non solo necessaria, ma divenne
un'attività così determinante, fondamentale e discriminante, quando i
nostri antenati potevano semplicemente camminare?
I dati archeologici testimoniano che dopo Australopithecus era
avvenuta negli ominidi un'importante e graduale transizione, che
aveva visto sempre più aumentare la componente carnivora nella dieta
dei nostri antenati. Il passaggio da un'alimentazione a base
vegetale a un'alimentazione a base animale comporta però cambiamenti
a catena, a causa ed effetto, non solo nelle abitudini alimentari e
comportamentali ma anche a livello scheletrico e anatomico. Le
piante sono infatti statiche, sessili, e sono dotate solo di
meccanismi di difesa passivi, quali spine, sostanze chimiche di gusto
repellente, fitoliti, o particolari abitudini di crescita. Gli
alimenti vegetali hanno generalmente un basso contenuto proteico e
un alto contenuto di fibre con un basso valore calorico. Hanno
inoltre un elevato contenuto di cellulosa indigeribile e lo stomaco e
l'intestino negli erbivori assumono quindi la funzione di rumine,
una sorta di "camera di fermentazione" dove viene scomposta la
cellulosa per liberarne le proprietà nutritive. Per gli erbivori il
tempo speso nella nutrizione, in relazione al peso corporeo,
raggiunge valori molto alti e un erbivoro impiega più tempo a
nutrirsi rispetto a un carnivoro dello stesso peso. Per un animale
di circa 3040 kg, come un Australopithecus, passare da una dieta
erbivora a una dieta carnivora significherebbe quindi spendere
invece di 6 solo 2 ore al giorno nella nutrizione, con un guadagno
di 4 ore, ovvero un terzo delle ore di luce all'equatore. La carne è
infatti prontamente digeribile, ha bisogno di un intestino
relativamente più breve rispetto agli erbivori, con una prevalenza di
intestino tenue e inoltre ogni preda rappresenta una grande fonte di
calorie, contenendo un alto numero di proteine e un basso numero di
fibre. Se però adottare una dieta a base di carne sembra essere
vantaggioso, l'approvvigionamento del cibo diventa cosa complicata. Gli
animali da preda sono infatti circospetti, mobili, e usano non solo
difese passive, quali la mimetizzazione, ma anche difese attive,
quali morsi, calci, e la fuga. Passare quindi a una dieta carnivora
richiede necessariamente una specializzazione
nell'approvvigionamento di carne, midollo e cervello, attraverso lo
sciacallaggio o attraverso la caccia, trovandosi così costretti a
competere con gli altri predatori della savana. Prima di riuscire a
costruire utensili, quali lance, frecce, reti (non testimoniate
prima del Paleolitico Superiore, quindi non prima di 40.000 anni
fa), capaci di bloccare e di colpire a lunga distanza la preda,
probabilmente la corsa rappresentava l'unica risorsa non solo per
sfuggire ai predatori, ma anche per entrare in possesso del cibo.
Anche solo per poter raggiungere e trasportare le carcasse prima di
altri animali è necessario aver sviluppato caratteristiche fisiche di
robustezza, resistenza, stabilità e velocità, ed è stato ipotizzato
che i nostri antenati potessero, grazie alla loro attitudine alla
corsa, inseguire sino a sfiancare le prede, per poterle così
catturare e uccidere o per potersi avvicinare a sufficienza da
scagliare pietre e bastoni, come è stato osservato per i Boscimani
dell'Africa del Sud. Certo una tale tecnica, in relazione alle
possibilità di successo, implica un grande dispendio di energie e di
tempo, ma comunque non considerare la corsa come parte integrante
nell'approvvigionamento del cibo sarebbe ugualmente azzardato.
L'uomo d'altro canto non è mai stato visto come un gran
corridore, e non è effettivamente capace di raggiungere alte velocità
(il massimo è di circa 43 km/h nei migliori atleti, contro i quasi
100 km/h di antilopi e i 110 km/h del ghepardo) ma, sulle lunghe
distanze, dimostra di essere competitivo. Innanzitutto bisogna
considerare che paradossalmente nella caccia non è necessario che il
predatore abbia una velocità massima più elevata della preda. Il
paradosso può essere spiegato pensando alla differenza tra la velocità
massima media di tutta una specie e la velocità raggiunta da un
singolo predatore durante un particolare inseguimento. Accanto a
questo inoltre va aggiunto che oltre la velocità massima va preso in
considerazione il tempo durante il quale tale velocità sarà mantenuta
costante. L'uomo può regolarmente correre per 10 km e anche per
distanze notevolmente maggiori (basti pensare alle maratone di 42 km
a cui ogni anno decine di migliaia di persone partecipano), a una
media di 19-25 km/h, distanze impensabili per qualsiasi altro
primate, e comparabili con altri quadrupedi specializzati nella
corsa, quali cani, lupi e iene, che percorrono rispettivamente 10,
14 e 19 km al giorno. Questo non significa che l'uomo possa
automaticamente battere sulle lunghe distanze mammiferi
specializzati, ma rimane comunque un corridore competitivo, a
differenza degli altri Primati ancestrali e attuali, capaci solo di
brevi, se pur veloci, scatti. L'altro fattore in gioco nella caccia è
la socialità. I carnivori sociali, quali leoni e iene, hanno più
successo di quanto farebbe pensare il rapporto generale tra le
velocità del predatore e il buon esito dell'inseguimento, mentre i
predatori solitari, quali ghepardi, leopardi o le iene striate
africane, cacciano con esito peggiore. Anche solo cooperare per
isolare o per bloccare un animale da preda, o darsi il cambio nella
corsa, possono infatti diventare strategie vincenti nell'attività
venatoria.
I cambiamenti cruciali che quindi un predatore deve sviluppare
consistono in adattamenti anatomici al fine della resistenza e della
velocità, di modo da presentarsi competitivi, e adattamenti
comportamentali ai fini della socialità. Non è da sottovalutare
inoltre che con il passaggio a una dieta carnivora molti predatori
occupano il tempo in avanzo per dormire e per socializzare, e questo
potrebbe essere stato vero anche per i primi ominidi: forse proprio
il tempo libero impiegato nell'oziosa manipolazione di oggetti e
nella socializzazione rappresenta un carattere essenziale per lo
sviluppo dell'intelligenza e della creatività caratteristiche
dell'uomo moderno. Ma se da una parte il passaggio a una dieta
carnivora rappresenta la possibilità, o necessità, di dedicarsi ad
attività biologicamente stimolanti, quali la caccia e la
socializzazione, l'aumento del volume cerebrale richiede una maggiore
spesa di energia, e quindi un'alimentazione più ricca. Il cervello
consuma, rispetto al suo peso, il 3% delle riserve energetiche del
corpo, una quantità di energia cinque volte superiore a quella dei
muscoli, a prescindere dalla loro attività, e da solo necessita del
20% circa di tutto l'ossigeno che respiriamo. E' quindi sempre in un
meccanismo di causa-effetto che si dovrebbe leggere l'evoluzione
umana, non tanto cercando di stabilire quali delle acquisizioni sia
più importante, ma tenendo ben presente che, se pur in vario modo,
sono tra loro strettamente collegate e non scindibili l'una
dall'altra.
Contestualizzazione cronologica della corsa
Se è stato finora ipotizzato come e perché si sviluppò
l'adattamento alla corsa di resistenza, difficile resta stabilire
quando. E' possibile semmai ipotizzare quando ormai tale adattamento
fosse già sviluppato. Gli studi di Bramble e Lieberman delineano
infatti l'Homo come un corridore, una sorta di irrequieto viaggiatore,
che a piedi o a corsa si spostava spinto dalle necessità. Guardando
però alla nostra storia evolutiva il primo vero "viaggiatore" che
incontriamo è l'Homo ergaster mentre tutte le specie di ominidi
precedenti erano restate confinate all'interno del continente
africano. La prima grande diffusione di Homo è primato di ergaster. La
velocità con cui l'espansione territoriale avvenne è impressionante
considerando che la configurazione e il clima del territorio
cambiavano in continuazione, richiedendo una grande capacità
adattativa: dalla sua apparizione attorno a 1,8-1,7 Ma avrebbe in
breve tempo colonizzato la sponda orientale del Mediterraneo, almeno
da quanto testimoniato dai resti fossili di Homo georgicus rinvenuti a
Dmanisi in Georgia e databili attorno a 1,7-1,8 MA (i fossili
relativi ad H. georgicus, se pur attribuiti provvisoriamente alla
specie H. ergaster, potrebbero rappresentare una prima migrazione di
una forma arcaica differenziatasi in Africa prima della separazione
di H. ergaster, presentando difatti caratteri ancora molto vicini
ad H.
habilis, quali una capacità cranica ridotta, attorno a 600 cc, e
una cultura materiale ancora legata alla tradizione Olduwaiana),
per poi occupare il Vicino Oriente e l'Asia già attorno a 1,7-1,6
MA. Bisogna pensare che un'espansione tanto grande in un periodo
così breve di tempo sia da imputare anche all'affermato cambiamento
di dieta, essendo gli erbivori più legati a un clima e un ambiente
particolare, ed essendo inoltre necessario per erbivori adattati a
carnivori ridurre la propria densità demografica, probabilmente
proprio distribuendo la popolazione in una zona geografica più vasta. I
resti fossili testimoniano che H. ergaster era effettivamente del
tutto adattato a una dieta carnivora: possedeva infatti un intestino
breve, caratteristica esclusiva dei carnivori, venendo le proteine
animali metabolizzate velocemente a differenza degli alimenti
vegetali. Il torace dell'ergaster era infatti cilindrico,
particolarmente lungo e stretto, quasi indistinguibile da quello
moderno, e sicuramente non poteva ospitare un intestino lungo come
quello necessitato dagli erbivori per trasformare il cibo.
H. ergaster era quindi carnivoro. Era anatomicamente molto
vicino all'uomo moderno, probabilmente cacciava, a giudicare dagli
strumenti in pietra che segnano un deciso balzo qualitativo rispetto
alle industrie Olduwaiane ed è anche attestato lo sviluppo di stretti
legami sociali tra individuo e individuo. Soprattutto percorreva
lunghe distanze. Inoltre, dei 26 caratteri analizzati dagli studiosi
americani, ben 11 sui 15 coinvolti esclusivamente nella corsa di
resistenza sono con sicurezza riscontrati in Homo ergaster, e altri 4
gli sono invece attributi con un margine di incertezza. Questo non
vuole dimostrare che l'attitudine alla corsa si sia sviluppata con
ergaster ma semmai induce a pensare che già fosse sviluppata in questa
forma ominide.
Difficile poi stabilire sino a che punto lo scheletro e
l'anatomia di H. habilis si fosse differenziata rispetto ai precedenti
ominidi e se già presentasse evidenti adattamenti a una prolungata
attività di corsa, vista anche l'incertezza con cui i fossili sono
attribuiti a tale entità tassonomica.
La corsa come adattamento alla caccia
L'uomo, come Kortlandt già affermava, è l'unico animale a
riunire in se l'abilità manuale del Vegetariano alla concentrazione e
alla scaltrezza del Carnivoro. Caratteristiche fondamentali dei
Primati arboricoli sono infatti la differenziazione morfologica e
funzionale tra arti posteriori e arti anteriori, la differenziazione
delle dita e l'acquisizione di una loro mobilità indipendente,
soprattutto per quanto concerne l'indice e il pollice. Se pur con le
dovute proporzioni e differenze tra specie e specie, gli arti
anteriori assumono fondamentalmente una funzione prensile e di
sostegno, mentre gli arti posteriori una funzione di appoggio.
L'eredità lasciata dalla vita arboricola ha permesso all'uomo
un'ulteriore differenziazione morfologica e funzionale degli arti,
sviluppando da una parte mani capaci di compiere operazioni complesse e
delicate, e dall'altra una base d'appoggio capace di sostenere
l'intero peso corporeo, di adattarsi al terreno e di compiere
movimenti complessi al fine di mantenere l'equilibrio. Potremmo dire
che quindi l'uomo è anatomicamente un compromesso tra l'abilità
manuale, le capacità visive e la mobilità del Vegetariano arboricolo e
la propensione alla caccia del Carnivoro terricolo.
Potrebbe essere stato il graduale adattamento a predatore ad
aver richiesto lo sviluppo di strutture anatomiche che coniugassero
resistenza e stabilità ad agilità e mobilità, che consentissero la
pratica di attività di estremo sforzo fisico, quali la corsa,
indipendentemente dal sostegno degli arti superiori, liberi così di
specializzarsi nella manipolazione dell'ambiente esterno, e in primis
nella produzione di strumenti necessari alla sopravvivenza. Correre
quanto camminare sono attività distintive dell'essere uomo,
acquisizioni decisive nella nostra storia evolutiva, effetto e causa
di adattamenti ugualmente importanti, tanto radicate in noi che
spesso è difficile rendersi conto della loro complessità, e dei
sottili equilibri necessari al bipedismo "potenzialmente
catastrofico" che ci caratterizza.
Problematiche
Nonostante sia ragionevole ipotizzare un importante ruolo
dell'attività di corsa perlomeno nell'evoluzione di alcune parti o di
alcuni particolari complessi anatomici umani, sorgono ovviamente
numerose problematiche che necessariamente devono essere prese in
considerazione.
Innanzitutto è sempre difficile stabilire il preciso
coinvolgimento di un particolare gruppo muscolare od osseo nel
compiere una determinata azione. Non c'è infatti sempre una sola
maniera di compiere l'azione, di "muoversi", soprattutto per quanto
riguarda la corsa dove la variabilità nella realizzazione del
movimento è molto forte da individuo a individuo, a seconda della
velocità e del terreno dove si pratica, ma soprattutto è difficile
stabilire e quantificare quanto una particolare struttura anatomica
possa essere fondamentale per un'azione e non per un'altra, per la
"corsa" e non per la "camminata". Stabilire che poi una particolare
conformazione sia evoluta esclusivamente per compiere un dato
movimento è ancor più difficile da dimostrare. Alcune critiche mosse
alle deduzioni di Bramble e Lieberman che, se posso permettermi,
mettono un accento un po' troppo forte sul ruolo della camminata e della
corsa nel processo evolutivo, ("it is reasonable to hypothesize
that Homo evolved to travel long distance by both walking and
running."), sottolineavano che le nostre gambe e le nostre braccia
possono essere considerate anche adattate per il nuoto, ma questo
non significa che la loro anatomia si sia evoluta necessariamente
per questo. E' ovviamente una provocazione, restando la corsa
comunque un'attività più prepotentemente necessaria e naturale per
l'uomo che il nuoto, ma comunque è un interessante spunto per
ribadire che le necessità che hanno spinto l'evoluzione e modificato
le nostre caratteristiche anatomiche sono sicuramente molteplici,
dal bisogno di possedere arti liberi per il trasporto e la
produzione di oggetti, al bisogno di adottare una locomozione che
offrisse una visuale più ampia ed esponesse una maggiore superficie
corporea al vento e alle correnti d'aria fresca, soprattutto
considerando l'ambiente della savana.
Un'altra interessantissima critica è stata mossa da Patricia
Kramer dell'Università di Washington. Se la corsa di resistenza, si
domanda, è un'attività di competenza maschile, in quanto da legare
prevalentemente alla caccia e all'approvvigionamento del cibo, come
mai anche le donne presentano un bacino stretto e un grande gluteo
completamente sviluppato, e sono totalmente adattate alla corsa di
resistenza? Non è facile rispondere a un tale domanda, ma si possono
comunque fare alcune osservazioni. Innanzitutto la suddivisione dei
compiti che avrebbe visto protagonisti uomini e donne nei primi
aggregati umani non è un processo cronologicamente ben databile né
tanto meno è facile stabilire i modi e le tappe attraverso le quali si
realizzò. Si ipotizza che in tali gruppi, successivamente al
cambiamento di habitat e all'acquisizione della stazione eretta, le
femmine gravide o con piccoli erano difficilmente in grado di
partecipare alla caccia, cosicché venne a crearsi naturalmente una
separazione dei compiti. Solo le femmine senza piccoli e i maschi
prendevano parte alla caccia, le altre rimanevano in luoghi ritenuti
tranquilli e sicuri e si occupavano dell'allevamento dei piccoli,
della raccolta di frutti e altri alimenti. A poco a poco le femmine,
anche a causa di ripetute gravidanze, avrebbero però perso
l'allenamento alla caccia portando così a una più netta separazione dei
compiti e all'accumulo di maggiori differenziazioni anatomiche.
Ora, se verosimilmente questo può essere stato in breve il
processo avvenuto, bisogna comunque considerare che se pur
nell'attività di caccia è fondamentale la capacità di correre a buone
velocità anche per lunghe distanze, il processo di diversificazione
dei compiti può essere avvenuto in un periodo successivo, quando
l'attività di caccia, unita al maggiore sviluppo celebrale, alla
produzione di strumenti maggiormente elaborati in legno, osso e pietra
e allo sviluppo di mezzi di comunicazione sempre più specifici,
sarebbe divenuta sempre più complessa, elaborando vere e proprie
strategie d'azione, grazie a trappole e armi da lancio e richiedendo
una pratica costante e una partecipazione continua, sia per
apprendere le tecniche di caccia stesse, sia per riuscire a
coordinarsi efficientemente con gli altri partecipanti. La capacità
di correre su lunghe distante ad andature sostenute avrebbe
rappresentato invece un notevole vantaggio quando ancora tali
tecniche non si erano probabilmente evolute, non prima di 80.000
anni fa quindi, e quando lo sciacallaggio era ancora uno dei modi
principali di procurarsi cibo, rendendosi necessario competere con
altri mammiferi corridori senza disporre che di pietre più o meno
lavorate e bastoni. In questo quadro è possibile che non si fosse
ancora delineata una precisa suddivisione dei compiti, e che quindi
le femmine non fossero completamente escluse dall'attività
venatoria.
Accanto a questo bisogna considerare che comunque la corsa è
un'attività naturale, istintiva per l'uomo, da considerare anche come
un'attività di "difesa", un modo per fuggire dai predatori, per
percorrere velocemente tratti di savana in cui si è maggiormente
esposti all'azione di altri carnivori, un modo anche per poter
trasportare, nel caso si renda necessario, i piccoli inermi nel minor
tempo possibile e quindi esponendosi meno a eventuali pericoli.
Se pur sono necessari studi molto più approfonditi, anche in tal
caso probabilmente le acquisizioni umane sono da considerarsi come
prodotto di più necessità e di più componenti diverse, e per
un'attività come la corsa ipotizzare una differenziazione a seconda
dei sessi diventa, se pur molto interessante, molto rischioso e
problematico.